|
It has been a while since my last post. I needed to take some personal time and had to step away from science for a bit. After going through my first year at a postdoc, applying for faculty jobs (unsuccessfully), trying to finish up PhD projects, issues with intellectual and data sharing from colleagues, and some family emergencies along the way, I really needed to step back and focus on myself before I could come back to work. So apologies for the delay for those who follow these posts. I will be back on schedule from now on!
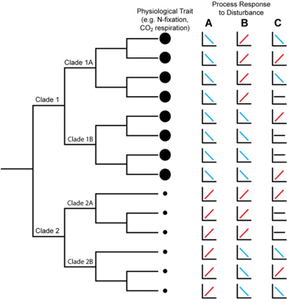
Framework to link microbial structure to function
Quantifying physiological traits What other endorsement do you need to read this next paper [3]. Understanding carbon use efficiency has wide-ranging applications from breaking down growth rate - yield tradeoffs to scaling CUE for ecosystem modeling. Using 23 diverse soil isolates, the authors investigated physiological traits, including growth rate on various substrates. By computing CUE metrics for each strain the authors could correlate these values to genomic predictions, such as rRNA operon copy number and extracellular enzymes. Some results included wide ranges in CUE temperature sensitivity across taxa, creating difficulties in predicting phylogenetic conservatism of an assumed complex trait. Brief note, I similarly found a wide range in carbon use across strains within a single genus and this variation was highly dependent on temperature [4] - what drives these differences?! Further, the authors found that genomic predicted traits did not correlate with CUE measurements, highlighting the difficulty in using genomic traits as reliable predictors. We always need to integrate physiology! Scaling from traits to response to whole communitiesThe best application of trait-based approaches in naturally occurring bacteria are probably the phytoplankton (specifically Prochlorococcus and Synechococcus). A new paper analyzed the distribution of picoeukaryotic phytoplankton with a global environmental abundance dataset combined with quantitative niche modeling [5]. The integration of the niche model enabled the authors to provide finer-taxonomic resolution to capture the high diversity of functional species within the phytoplankton groups (including differential responses to temperature and light). Together, the authors observed niche partitioning along a temperature gradient across the three major groups, which account for most of the photosynthetic biomass in tropical waters. Using metagenomics, microbial ecologists have gained an unprecedented insight into the functional potential of understudied taxa as well as whole communities. However, as the above paper suggests, the genomic potential doesn't always tell the entire story. This next paper utilizes time-resolved metatranscriptomics to understand how expressed genes in microbial communities contribute to functional observations [6]. By comparing metatranscriptomes across rhizosphere and bulk soil, the authors found a significant successional pattern across time. Further, functional genes indicated a rapid functional successional pattern as well with four major functional guilds emerging: specialization to root exudates, decaying roots, root biomass, and later stage roots. These overall patterns suggest that specific microbial taxa specialize on various spatiotemporal strategies related to both habitat (root v soil) and time. As a disclaimer to those interested in using metatranscriptomics, they are very messy datasets. Honestly, they make metagenomes look easy... Here, the authors capitalized on a well-known system (just check the author list!) that has previously created a unique reference database for this specific system, including single-cell genomes (SAGs), isolate genomes, and stable isotope probing metagenomes. Last paper I want to include really capture why assessing the (active) functional genes can better resolve community patterns. In particular, does relic DNA (essentially leftover DNA from dead organisms) mask community patterns [7]? Using fine-scale spatial sampling combined with temporal sampling, the authors demonstrate that, unsurprisingly, spatial variation largely explains community patterns. This is not new as distance decay relationships were an early observation for the argument FOR microbial biogeography patterns. What was interesting is the temporal dynamics. The observed temporal dynamics was hindered by the presence of relic DNA. By accounting for relic DNA in the samples, the authors found the temporal signal to be more evident. Papers:1. Bardgett RD, Caruso T. (2020). Soil microbial community responses to climate extremes: resistance, resilience and transitions to alternative states. Phil. Trans. R. Soc. B
DOI: 10.1098/rstb.2019.0112 2. Allison SD, Martiny JBH. (2008). Resistance, resilience, and redundancy in microbial communities. PNAS DOI: 10.1073/pnas.0801925105 3. Pold G, Domeignoz-Horta LA, Morrison EW, Frey SD, Sistla SA, DeAngelis KM. (2020). Carbon use efficiency and its temperature sensitivity covary in soil bacteria. mBio DOI: 10.1128/mBio.02293-19 4. Chase AB, Gomez-Lunar Z, Lopez AE, Li J, Allison SD, Martiny AC, Martiny JBH. (2018). Emergence of soil bacterial ecotypes along a climate gradient. Environmental Microbiology DOI: 10.1111/1462-2920.14405 5. Flombaum P, Wang WL, Primeau FW, Martiny AC. (2020). Global picophytoplankton niche partitioning predicts overall positive response to ocean warming. Nature Geoscience DOI: 10.1038/s41561-019-0524-2 6. Nuccio EE, Star E, Karaoz U, Brodie EL, Zhou J, Tringe SG, Malmstrom RR, Woyke T, Banfield JF, Firestone MK, Pett-Ridge J. (2020). Niche differentiation is spatially and temporally regulated in the rhizosphere. ISME DOI: 10.1038/s41396-019-0582-x 7. Carini P, Delgado-Baquerizo M, Hinckley ELS, Holland-Moritz H, Brewer TE, Rue G, Vanderburgh C, McKnight D, Fierer N. (2020). Effects of spatial variability and relic DNA removal on the detection of temporal dynamics in soil microbial communities. mBio DOI: 10.1128/mBio.02776-19
1 Comment
|
AuthorSome thoughts on some (small) things Archives
May 2023
Categories |

 RSS Feed
RSS Feed